Part 1: Anatomy and Histology of the heart
Mon Jul 06 2026
By B. Hassan

As I get closer to finishing med school, I started to consider what specialty I want to pursuit. And considering all the specialties I did so far, noting caught my attention as much as cardiology. It’s a specialty in which diving deeper and reading more is both cognitively challenging and extremely rewarding. It’s also based on many concepts from physics, which I extremely enjoy, especially because it can help break the monotony of biology. Moreover, it’s clinically very rewarding, with long-term physician-patient relationships, and one of the specialties in with you can make a difference in both patient’s morbidity and mortality, especially alluding to how common mortality from cardiac causes is.
Just after making this decision, I started to look for avenues to revise my cardiology training throughout med school. One of the resources that stood out is the amazing book Pathophysiology of Heart disease by Leonard S. Lilly. While reading this book and taking notes, I though that sharing my notes can be extremely valuable for undergraduate med students or for beginner residency doctors who desire a quick review of cardiology topics. This is not a substitute for reading the book, and if you have the time and financial means to get the book, then it is highly recommended.
And now after this lengthy introduction, let’s start with the anatomy and histology of the heart.
Pericardium
Pericardium is a fibroserous sac that encloses the heart along with the roots of great vessels. This structure consists of two layers, a strong outer fibrous layer and an inner serosal layer.
The inner serosal layer adheres to the external wall of the heart and is called the visceral pericardium. The visceral pericardium reflects back on itself (like a sheet of paper that folds on itself) and lines the outer fibrous layer, forming the parietal pericardium.
The space between the visceral and parietal pericardial layers contains a thin film of pericardial fluid that acts as a lubricant, allowing the heart to beat in a minimal-friction environment.
The pericardium is attached to the sternum and mediastinal portions of the right and left pleurae. Its many connections to the surrounding structures keep the pericardial sac firmly anchored within the thorax.
Surface anatomy
The heart is cone-shaped with an apex that is formed by the tip of the left ventricle and a base or posterior surface that is formed by the atria, mainly the left.
- The apex points inferiorly, anteriorly, and to the left.
- The base or posterior surface lies between the lung hila.
The anterior surface of the heart is formed by the right atrium and ventricle. Because the left atrium and ventricle lie posteriorly, they form only a small strip of this anterior surface.
The inferior (diaphragmatic) surface of the heart is formed by both ventricles, primarily the left.
Heart borders
Observing the chest from an anteroposterior view, 4 borders of the heart are apparent.
- The right border is formed by the right atrium
- The left border is formed by the left ventricle and a portion of the left atrium
- The superior border is formed by both atria.
- The inferior border is formed mainly by the right ventricle, with a slight contribution from the left ventricle near the apex.
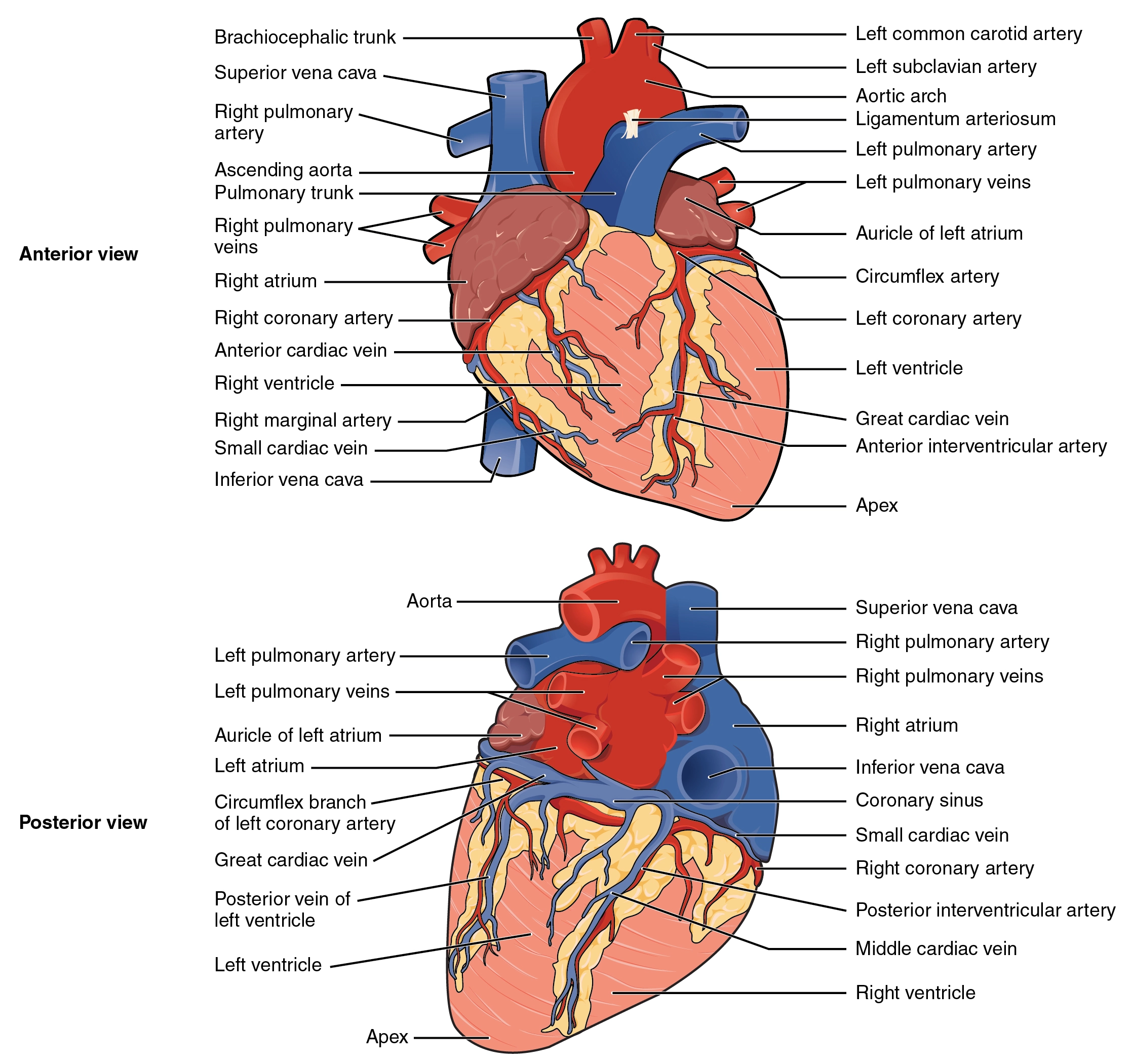
Internal structures
All four heart valves are attached to the fibrous cardiac skeleton, which is located between the atria and ventricles. The cardiac skeleton also serves as a site of attachment for the ventricular and atrial muscles.
The surface of the heart valves and the interior surface of the chambers are lined by a single layer of endothelial cells known as the endocardium. This is anatomically and functionally identical to the endothelium that forms the tunica interna (intima) of blood vessels. It provides the exact same smooth, non-thrombogenic surface to keep blood flowing smoothly without clotting.
The subendocardial tissue contains fibroblasts, elastic and collagenous fibers, veins, and mainly contain branches of the conducting system and is continuous with the myocardium.
The epicardium is the outermost layer of the heart and is just another term for the visceral layer of the serous pericardium.
The major vessels travel within the epicardial fat and then send branches that penetrate the myocardium. As they dive deeper, they branch into smaller and smaller arterioles and capillaries to supply the myocardium. They finally terminate in a dense plexus in the subendocardial tissue.
Each annulus fibrous surrounding the mitral and tricuspid valves is thicker than those surrounding the pulmonic and aortic valves.
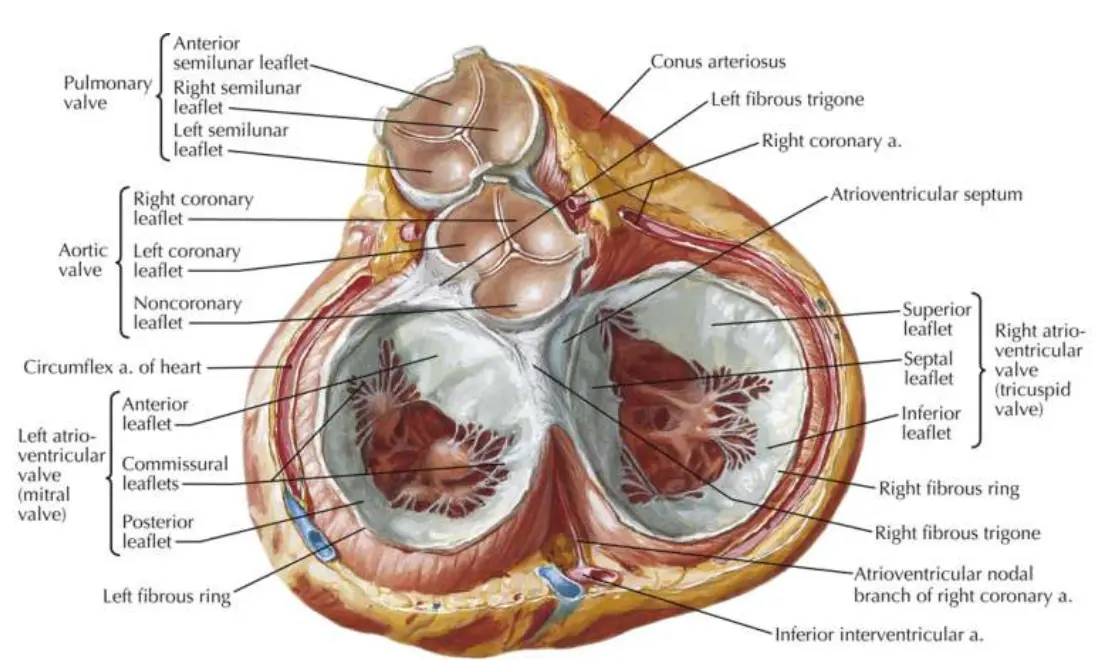
Right side
The right ventricle is roughly triangular in shape, and its superior aspect forms a cone-shaped outflow tract, which leads to the pulmonary artery. Although the inner wall of the outflow tract is smooth, the rest of the ventricle is covered by a number of irregular fleshy columns known as trabeculae carneae.
A large trabecula that crosses the ventricular cavity is called the moderator band. It carries a component of the right bundle branch of the conducting system to the ventricular muscle.
The right ventricle contains three papillary muscles, which project into the chamber and via their thin, stringlike chordae tendineae attach to the edges of the tricuspid valve leaflets. The leaflets, in turn, are attached to the annulus fibrous that supports the valve between the right atrium and ventricle.
Contraction of the papillary muscles prior to other regions of the ventricle tightens the chordae tendineae, helping align and restrain the leaflets of the tricuspid valve as they are forced closed. This prevents blood regurgitation into the right atrium during right ventricular contraction.
Left side
The cavity of the left ventricle is approximately cone shaped and longer than that of the right ventricle. In a healthy adult heart, the wall thickness is 9-11 mm, roughly 3 times that of the right ventricle.
The aortic vestibule is a smooth-walled part of the left ventricular cavity. Inferior to this region, most of the ventricle is covered by trabeculae carneae, which are finer and more numerous than those in the right ventricle.
The left ventricular chamber contains two large papillary muscles. These are larger than their counterparts in the right ventricle, and their chordae tendineae are thicker but less numerous. The chordae tendineae of each papillary muscle distribute to both leaflets of the mitral valve.
Similar to the right ventricle, tensing of the chordae tendineae during left ventricular contraction helps restrain and align the mitral leaflets, enabling them to close properly and preventing backward leakage of blood.
And similar to the right atrium, the leaflets of the mitral valve are attached to the annulus fibrous that supports the valve between the left atrium and left ventricle.
Just above the right and left aortic valve cusps in the aortic wall are the origins of the right and left coronary arteries.
Trabeculae Carneae
Trabeculae carneae are the coarse, irregular muscular ridges that line the inner walls of the ventricles. They look like thick, fleshy columns. Most of them are attached flat against the heart wall along their entire length, while some form free bridges anchored at both ends.
They are made of actual, contractile myocardium covered by endocardium.
Example: the moderator band in the right ventricle is technically a massive, specialized type of trabecula.
Note
Moderator band = septomarginal trabecula
Function
Trabeculae Carneae break up the smoothness of the ventricular walls. Think about what happens when you press two flat pieces of wet glass tightly together. They stick. To pull them apart, you have to fight against a powerful suction effect caused by surface tension and fluid adhesion.
Without Trabeculae Carneae, during systole, the ventricle squeezes incredibly hard to eject blood, and its walls come into direct contact with each other. If the walls were smooth, wet endocardial surfaces would flatten against one another and “glue” shut at the peak of the squeeze. During Diastole (Filling), the heart would have to waste a massive amount of energy just to peel its own walls apart before it could even begin to fill with blood again.
They also help reduce blood turbulence. If the ventricle walls were completely flat, blood rushing into the chamber during filling would slam against the walls, creating large, chaotic swirling whirlpools. This chaotic swirling wastes kinetic energy and makes the heart work harder to redirect that blood out into the aorta or pulmonary artery. This textured surface actually helps “channel” the blood smoothly along the wall, conserving its kinetic energy and cleanly swirling it toward the outflow tract so it can be ejected efficiently.
False Chordae Tendineae
“True” chordae tendineae are the string-like cords that connect papillary muscles to the valve leaflets to keep them from flipping backward.
False chordae tendineae look exactly like true chordae, but they don’t connect to the wrong places. Instead of anchoring a valve, they freely cross the ventricular cavity, connecting one wall to another, bridging the septum to a papillary muscle, or connecting two papillary muscles together.
They are primarily fibrous connective tissue, and they look like thin, white, stringy, tendon-like cords.
They have no role in the heart function, and they are just considered benign anatomical variants. However, because they cross open space like strings on a guitar, they can vibrate in the bloodstream and sometimes cause innocent (harmless) murmurs.
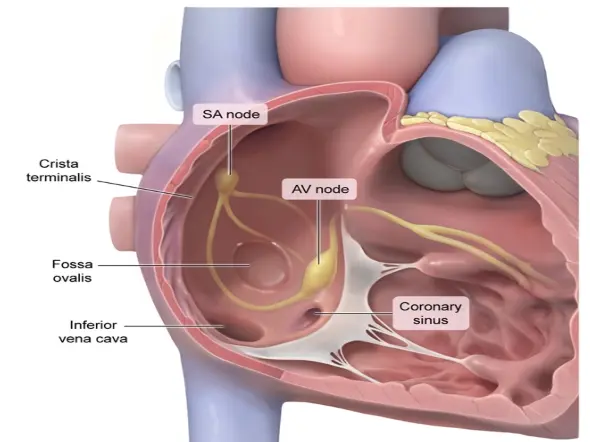
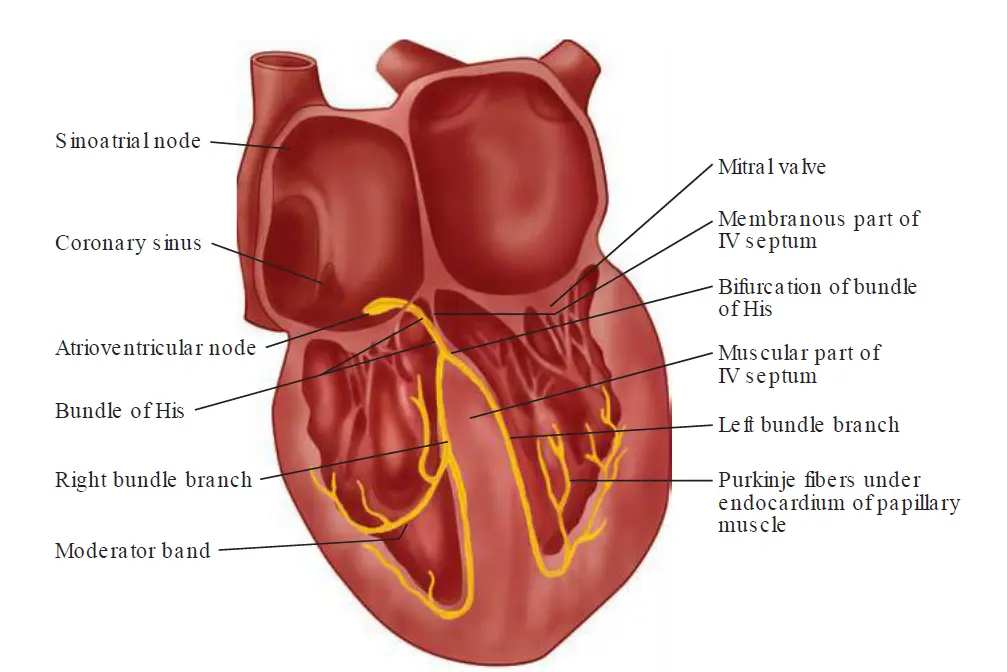
Electro-conductive system
The sinoatrial (SA) node is a small mass of specialized cardiac muscle fibers in the wall of the right atrium. It is located to the right of the superior vena cava entrance and normally initiates the electrical impulse.
The atrioventricular (AV) node lies just beneath the endocardium in the inferoposterior part of the interatrial septum.
Distal to the AV node is the bundle of His, which perforates the interventricular septum to bifurcate into a cablelike structure on the right side of the septum, known as the right bundle branch, and a broad sheet of fibers that continues over the left side known as the left bundle branch.
Near the junction of the interventricular septum and the anterior wall of the right ventricle, the right bundle branch becomes subendocardial and bifurcates. One branch travels across the right ventricular cavity in the moderator band, whereas the other continues toward the tip of the right ventricle. These branches eventually arborize into a finely divided anastomosing plexus that travels throughout the right ventricle.
Functionally, the left bundle branch is divided into an anterior and a posterior fascicle and a small branch to the septum. The anterior fascicle runs anteriorly toward the apex, forming a subendocardial plexus in the area of the anterior papillary muscle. The posterior fascicle travels to the area of the posterior papillary muscle; it then divides into a subendocardial plexus.
The subendocardial plexuses of both ventricles send distributing Purkinje fibers to the ventricular myocytes. Impulses within the His–Purkinje system are transmitted first to the papillary muscles and then throughout the walls of the ventricles, allowing papillary muscle contraction to precede that of the ventricles.
How exactly does the papillary muscles contract before the rest of the ventricle?
In the Right Ventricle: The electrical signal takes a massive shortcut through the moderator band (septomarginal trabecula), which carries the right bundle branch straight across the ventricular cavity directly to the base of the anterior papillary muscle.
In the Left Ventricle: The left bundle branch divides into fascicles. These two fascicles act as direct, uninterrupted express pipelines shooting straight down the interventricular septum. Their primary, direct targets are the bases of the two papillary muscles. The main wave of electricity hits the papillary muscles first because they are the physical destinations of these major electrical trunks. Only after these thick fascicles reach the papillary muscles do they explode into the finer, web-like network of individual Purkinje fibers that spread across the rest of the left ventricular wall.
Cardiac vessels
The heart muscle is supplied by the right and left coronary arteries, which arise from the root of the aorta just above the aortic valve cusps. After their origin, these vessels pass anteriorly, one on each side of the pulmonary artery.
The large left main coronary artery passes between the left atrium and the pulmonary trunk to reach the AV groove. There it divides into the left anterior descending (LAD) coronary artery and the circumflex artery.
The LAD travels within the anterior interventricular groove toward the heart apex. During its descent, it gives off septal branches that supply the anterior two thirds of the interventricular septum and the apical portion of the anterior papillary muscle. The LAD also gives off diagonal branches that supply the anterior surface of the left ventricle.
The circumflex artery continues within the left AV groove and passes around the left border of the heart to reach the posterior surface. It gives off large obtuse marginal branches that supply the lateral and posterior wall of the left ventricle.
The right coronary artery (RCA) travels in the right AV groove, passing posteriorly between the right atrium and ventricle. It supplies blood to the right ventricle via acute marginal branches. In most people (roughly 85% of people), the distal RCA gives a large branch, the posterior descending artery. This vessel travels from the inferoposterior aspect of the heart to the apex and supplies blood to the inferior and posterior walls of the ventricles and the posterior one third of the interventricular septum. Just before giving off the posterior descending branch, the RCA usually gives off the AV nodal artery.
When the posterior descending and AV nodal arteries arise from the RCA, this is known as right dominant coronary circulation. In approximately 8% of the population, the posterior descending artery arises from the circumflex artery instead, resulting in a left dominant circulation. In the remaining population, the heart’s posterior blood supply is contributed to from branches of both the RCA and the circumflex, forming a codominant circulation.
The blood supply to the SA node is also most often (70% of the time) derived from the RCA. However, in 25% of normal hearts, the SA nodal artery arises from the circumflex artery, and in 5% of cases, both the RCA and the circumflex artery contribute to this vessel.
From their epicardial locations, the coronary arteries send perforating branches into the ventricular muscle, which form a richly anastomosing vasculature in the walls of all cardiac chambers.
Collateral connections, usually less than 200 µm in diameter, exist at the subarteriolar level between the coronary arteries. In the normal heart, few of these collateral vessels are visible. However, they may become larger and functional when atherosclerotic disease obstructs a coronary artery, thereby providing blood flow to distal portions of the vessel from a non-obstructed neighbor.
Coronary veins
The coronary veins follow a distribution similar to that of the major coronary arteries. These vessels return blood from the myocardial capillaries to the right atrium predominantly via the coronary sinus. The major veins lie in the epicardial fat, usually superfcial to their arterial counterparts.
The thebesian veins provide an additional potential route for a small amount of direct blood return to the cardiac chambers.
thebesian veins
The muscle fibers located just beneath the endocardium are supplied either by the terminal branches of the coronary arteries or directly from the ventricular cavity through tiny vascular channels, known as thebesian veins, or in modern anatomy known as smallest cardiac veins.
While they can be found in all four chambers of the heart, they are most abundant in the right atrium and the right ventricle, and are least common in the left ventricle.
Flow of blood in thebesian veins
Because Thebesian veins are valveless, blood simply moves down its pressure gradient from high pressure to low pressure.
During systole, the contracting muscle fibers create an immense intramyocardial pressure () within the heart wall. This pressure is actually higher than the pressure inside the ventricular cavity (), especially in the deep subendocardial layers. The blood trapped inside the microscopic Thebesian capillaries and venules is violently squeezed out of the wall. Because , the blood is milked forward, squirting out of the Thebesian foramina directly into the ventricular chambers.
During diastole, the ventricular muscle relaxes completely so the chamber can fill with blood from the atria. The intramyocardial pressure () drops almost to 0 as the muscle relaxes. Meanwhile, the pressure inside the ventricular chamber () rises as it fills with blood. Now, the gradient is reversed: . Because these tiny veins lack valves to block backward flow, the higher pressure inside the filling chamber can actually push blood backward into the Thebesian openings, filling the tiny vessels within the relaxed wall. This retrograde movement of blood into thebesian veins help supply the subendocardial tissue during diastole.
Note
The Thebesian veins dump deoxygenated blood from the heart muscle directly into the freshly oxygenated blood that is about to be pumped out to the body. Because of this constant trickle of deoxygenated blood from the left-sided Thebesian veins (alongside the bronchial veins from the lungs), the blood in the systemic arteries is never quite 100% saturated with oxygen, it typically sits around 95% to 98%.
The right ventricle only pumps to the lungs, so its maximum systolic pressure is quite low (around ). Because the chamber pressure is low, the Thebesian veins can easily drain blood into the right ventricle throughout almost the entire cardiac cycle. It acts as a highly efficient, functional drainage pathway.
The left ventricle pumps to the entire body, sky-rocketing to during systole.
- Because the crushing forces in the left ventricular wall are so extreme, the Thebesian veins here are completely obliterated and flattened during systole.
- During diastole, the left ventricular tissue relies almost entirely on the high-pressure coronary arteries to push fresh blood into the muscle from the outside-in.
This is why Thebesian veins are abundant and highly functional in the right atrium and right ventricle, but are largely minimized and compressed to the point of near-non-functionality in a healthy, normal left ventricle.
Histology
The cardiac myocytes measures up to 25 μm in diameter and 100 μm in length. The cell shows a cross-striated banding pattern similar to that of skeletal muscles. However, unlike the multinucleated skeletal myofibers, myocardial cells contain only one or two centrally located nuclei. Surrounding each myocardial cell is connective tissue with a rich capillary network.
Each myocardial cell contains numerous myofibrils, which are long chains of sarcomeres, the fundamental contractile units of the myocyte. Each sarcomere is made up of two groups of overlapping filaments of contractile proteins, actin and myosin. Biochemical interactions between the actin and myosin produce muscle contraction.
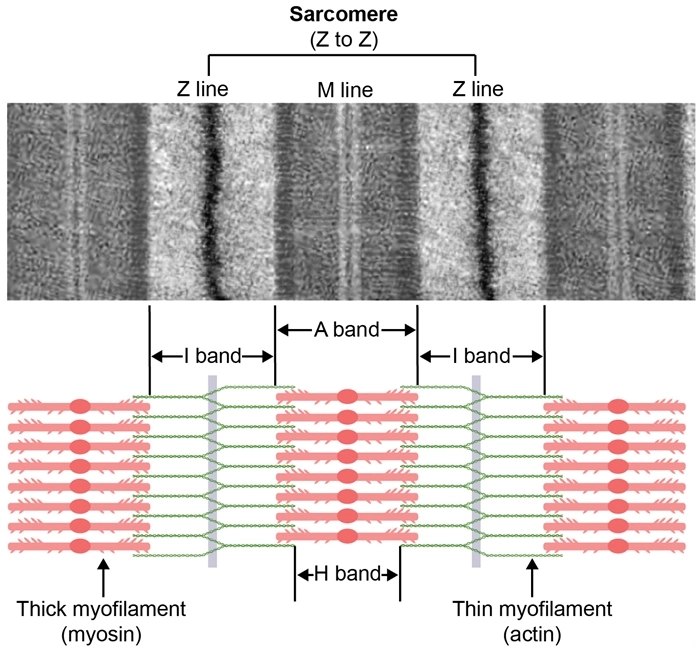
Under physiologic conditions, the overall sarcomere length (Z-to-Z distance) varies from 2.2 to 1.5 μm during the cardiac cycle. The larger dimension reflects fiber stretch during ventricular filling, whereas the smaller dimension represents the extent of fiber shortening during contraction
The myocardial cell membrane is called the sarcolemma. A specialized region of the membrane is the intercalated disk, a distinct characteristic of cardiac muscle tissue. Intercalated disks represent the gap junction complexes between adjacent cardiac fibers, which establish electrical continuity between the myocardial cells, allowing the whole ventricle to contract as one unit.
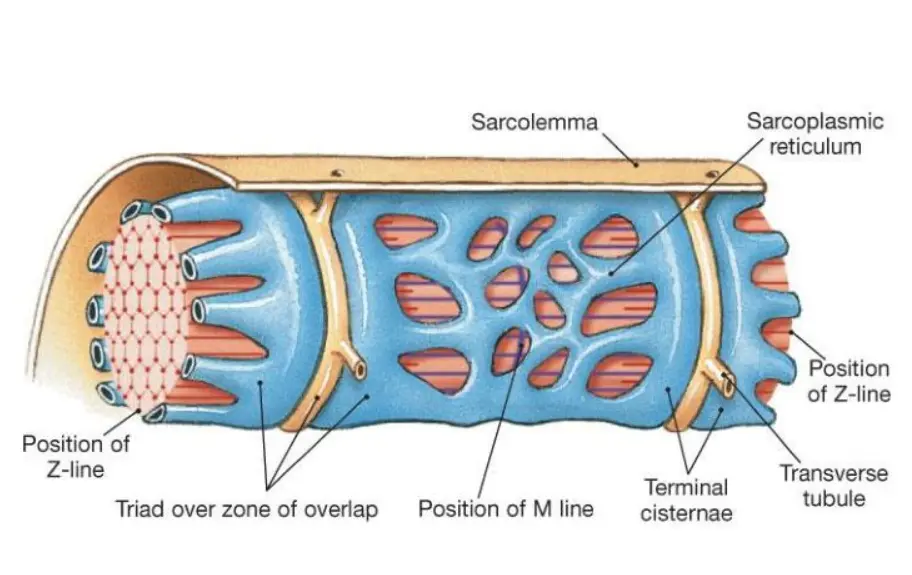
Another feature of the cell membrane is the transverse tubular system or T tubules. This is characterized by deep, fingerlike invaginations of the sarcolemma along the myofibrils. Similar to the intercalated disks, T tubules establish pathways of rapid transmission of the electrical impulses that initiate contraction.
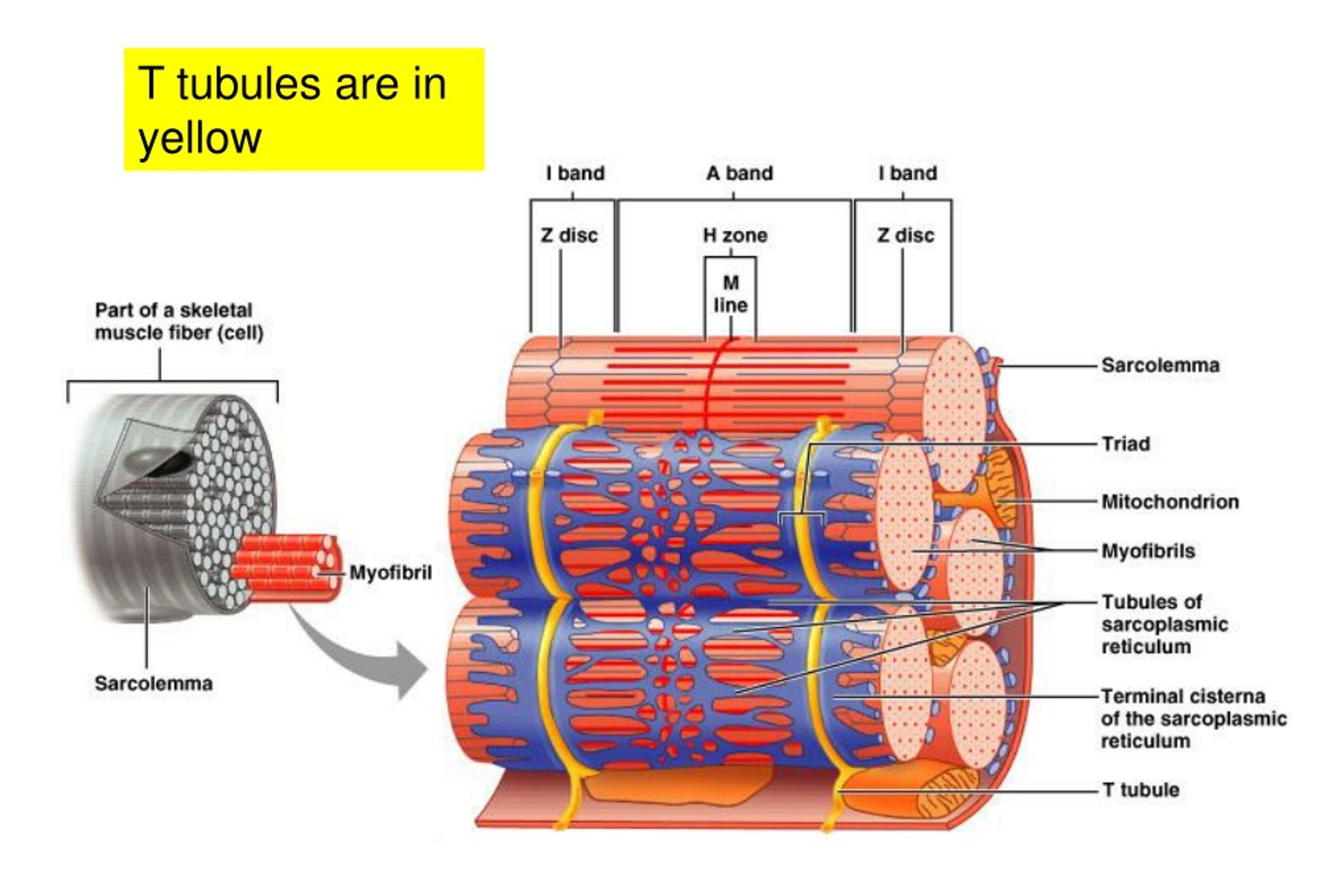
The sarcoplasmic reticulum (SR) is the myocyte analog of the endoplasmic reticulum. It is an extensive intracellular tubular network that complements the T tubule system. The SR intersects the T tubules in sacs called the terminal cisternae. These sacs house most of the intracellular calcium stores; the release of these stores is important for initiation of muscle contraction. Terminal cisternae also intersect the intercalated disks and the sarcolemma, providing each with a complete system for excitation–contraction coupling.
To serve the tremendous metabolic demand of the heart, the myocardial cell has an abundant concentration of mitochondria located between the individual myofibrils. These constitute approximately 35% of cell volume.